

Analysis: How Did the Tree of Life Grow Its Branches?
By Leonardo Campagna
Oilbird by Dusan Brinkhuizen/Macaulay Library. June 25, 2021More From Living Bird
 Living Bird Summer 2021—Table of Contents
Living Bird Summer 2021—Table of Contents Living Bird Magazine—Latest Issue
Living Bird Magazine—Latest Issue Living Bird Magazine Archives
Living Bird Magazine Archives
From the Summer 2021 issue of Living Bird magazine. Subscribe now.
There are some strange birds out there.
The Oilbird, for example, is a fruit-eating, cave-nesting bird from northern South America that gets its name from the use humans found for it in the past (using the bird’s fat as a source of oil to burn in lamps). Oilbirds are highly unusual nocturnal birds that can echolocate like bats, by emitting high-frequency clicks and listening for the returning echo. The Hoatzin is another weird one from South America, a leaf-eating bird the size of a small turkey. Hoatzins are also known as “stink-birds” because of the smell produced by fermenting leaves in their throat, similar to the way a cow digests food in its rumen.
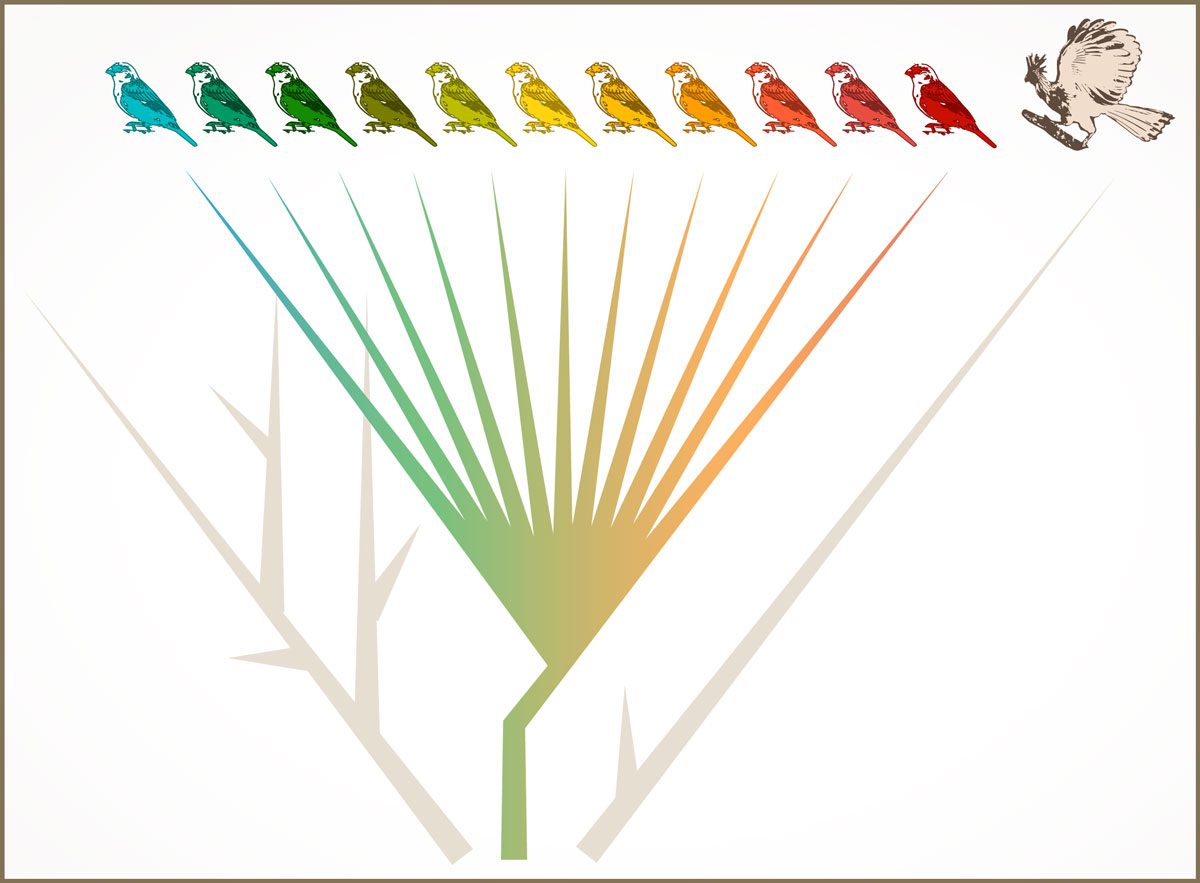
These two species are not only weird, but also unique from an evolutionary perspective. Neither Oilbird nor Hoatzin is closely related to any other living bird (see Add Flair to Your Life List by Tracking Down Evolution’s Most Distinctive Birds, Autumn 2018). On the bird tree of life they are each found on a solitary, long branch. Elsewhere on the bird tree there are large and closely related clusters of species, often referred to as evolutionary radiations. Darwin’s finches from the Galápagos Islands are the most famous example of a radiation, with many species generated in a comparatively short period of time.
The contrast between the Oilbird and Darwin’s finches highlights a key question in evolutionary biology: What makes certain groups of species more prone to multiply than others? Why are there so many warblers and so few swans? As an avian evolutionary biologist, I study this question via a group of small South American songbirds known as capuchino seedeaters. This cluster of 12 closely related seedeaters, which differ just slightly in the way males look and sound, represents the different ways capuchinos attract mates and scare off competitors.
Read More
 By Shuffling Genes, a Songbird Takes a Shortcut to Speciation
By Shuffling Genes, a Songbird Takes a Shortcut to Speciation How Can 9 Species Look So Different Yet Be Genetically Almost Identical?
How Can 9 Species Look So Different Yet Be Genetically Almost Identical? Add Flair to Your Life List By Tracking Down Evolution’s Most Distinctive Birds
Add Flair to Your Life List By Tracking Down Evolution’s Most Distinctive Birds
The key to understanding the capuchino seedeater cluster is in their genes. These birds are 99% identical, but the few areas of their genomes where they differ include genes that control plumage coloration. Much of the variation in male color patterns likely arises through hybridization. When on occasion different capuchino species interbreed, their genetic material is mixed and remixed.

Hybridization seems to provide a faster path to speciation—in the case of seedeaters, producing a unique new male color pattern that signals the beginning of a new species—than genetic mutation. Most species begin as products of isolation and time, the result of gene pools separated by a physical barrier (like an ocean, a desert, or a mountain). The pace of this process is dictated by the pace of random and infrequent mutations, so it is essentially a waiting game.
Speciation through the sharing of genetic material, as with seedeaters, saves time, because evolution can test a new trait in one species and then share it with a second species, generating something new almost instantly. Not all species can hybridize. This is why so many seedeaters were generated over perhaps a few hundred thousand years, whereas the Hoatzin has been on its own for millions of years. Scientists think the Hoatzin originated in Africa and rafted over on vegetation islands to South America. There it’s been for maybe an epoch or two, with no mutations as yet arising among isolated populations to give us a new species of Hoatzin in the Amazon that has survived to the present day.

There are other areas on the bird tree of life that scientists have yet to resolve, called polytomies. These are areas of the evolutionary tree where scientists haven’t yet figured out the relationships among several species. One particularly notable polytomy in the early branches of the bird tree, deep in the past, contains many large groups like waterbirds and shorebirds (although there are contested hypotheses on how these groups are related). We used to think solving these polytomies was a matter of adding more data, or using better mathematical algorithms and faster computers. But maybe these deep splits in the bird tree began like the capuchinos, with several similar species produced by the sharing of genetic material.
The bird tree could have started like an evolutionary shrub, with tangled and interconnected branches that got pruned by time, random chance, competition, and evolution. Some branches were pruned lightly, while others were pruned all the way to a single solitary branch—where echolocating Oilbirds and leaf-fermenting Hoatzins persist.

All About Birds
is a free resource
Available for everyone,
funded by donors like you
American Kestrel by Blair Dudeck / Macaulay Library