

An Enigma, Wrapped in a Riddle, Inside a Pine Cone?
By Jack Connor
April 15, 2009




Ask birders to name the toughest identification problems in North America, and you will probably hear about the spotted thrushes, several western hummingbirds, two or three genera of flycatchers, Cooper’s vs. Sharp-shinned hawk, Western vs. Semipalmated sandpiper, and a handful of other well-known puzzles.
Given a close enough look or listen, however, the sharpest observers can name most individuals in any of those groups. The field marks are subtle but generally reliable—and they are not the only clues. In fact, given the right habitat, timing, and geographic circumstances, even birders of modest abilities can identify most birds in all of those tough groups. Show me a mid-sized adult accipiter in South Carolina in June, and I will tell you, “That’s a Cooper’s, obviously”; show me one in Alaska in June, I’ll tell you, “That’s a no-doubt-about-it sharpie.” Breeding range is the key, of course. Not many closely related, look-alike species overlap on their nesting grounds, and even when they do, differences in foraging preferences, habitats, and seasonality usually help us separate one from the other.
Factoring geography and habitat into an ID may seem like cheating, but all experienced birders do so regularly, and basic biology supports the practice. According to classical Darwinian thinking, geographic isolation and differing habitats have created the planet’s biodiversity. Together with natural selection, they drive adaptive radiation, racial variation, and given enough time, create new species.
The finches on the Galápagos are the textbook example, of course. Apparently, a few individuals of a single finch species wandered to the archipelago several hundred miles off the coast of South America and survived there to breed, isolated from their relatives on the mainland. Natural selection, operating in a very different habitat than their homeland, changed them over time. Their descendents eventually drifted to other islands and may also have been joined by later colonists from the mainland. Because each island held different habitats and a different set of resident competitors, the finches were modified still more. After a few hundred thousand years of wandering and selection, 14 different species of Darwin’s finches now occupy those islands, each of them a more-or-less closed system of interbreeding individuals (hybrids occasionally occur).
For North American examples of this arrangement, think of the Island Scrub-Jays on Santa Cruz Island, isolated from their closest relatives (Western Scrub-Jays) by a dozen miles of Pacific Ocean off Santa Barbara, California, or the “Ipswich” Savannah Sparrows on Sable Island, isolated from mainland Savannah Sparrows by 100 miles of Atlantic Ocean off Nova Scotia, Canada. The first is currently considered a full species, the second a subspecies, but in both cases geography and nature have created a new set of creatures by separating them from their ancestral nesting grounds and reshaping them in response to their new environment.
So, how does this relate to North America’s toughest identification puzzle—the Red Crossbill?
Traditionally considered a single species of multiple races, Loxia curvirostra may actually comprise more than half a dozen separate species—each of them showing a mix of traits so close to the others that they are inseparable in the field. Even taxonomists with specimens in the hand are often uncertain of what they have. Birds of different bill sizes and shapes apparently prefer different conifer species (the smallest types feeding on hemlocks, the largest on white pines), but the differences must be measured in millimeters, and all groups feed on the seeds of different trees when their primary preferences are unavailable. Still more perplexing, all populations are nomadic and apparently able to nest anywhere in their widely varying ranges, at any time, from early winter through late summer, whenever local seed crops are rich enough. No birder should feel certain making an in-the-field ID of any member of this group based on geography, seasonality, foraging behavior, body size, bill shape, or plumage.
Taxonomists cannot even agree about what we should call the several populations. Are they a super-species? Sibling species? Subspecies?
Related Stories


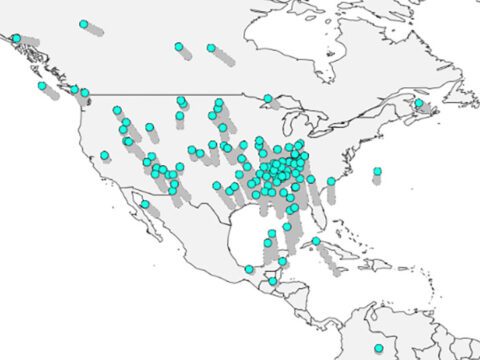

Ludlow Griscom called them subspecies in his landmark 1937 analysis, “A Monographic Study of the Red Crossbill.” He described eight different populations, distinguished by slight differences in body size, bill size, bill shape, and apparently differing breeding areas. Using the scattered information available at the time, he noted that each subspecies seemed to have a normal home range, although they sometimes bred outside those ranges. He was applying standard Darwinian thinking here: by definition, a subspecies can only be maintained by geographic isolation. The population is marked by racial differences, but it is not yet fully distinct from its closest relatives. If the breakaway group and the ancestral group breed separately long enough, they can become so different that they no longer consider each other potential mates if they encounter each other. Then, they are independent species. But first they must pass through an extensive period of isolation in which each is a subspecies—one breeding in one place, the other elsewhere. If they rejoin too soon, males and females of the two populations will merge their genes and the racial differences will disappear.
So, if the crossbill populations are subspecies, they should not overlap on their nesting grounds. Griscom had evidence that they did overlap: at least four of the subspecies he named—“Bendire’s crossbill,” L. c. bendirei; the “Sitka crossbill,” L. c. sitkensis; “Grinnell’s crossbill,” L. c. grinelli; and the “Mexican crossbill,” L. c. stricklandi—apparently nested in California, for example. But he got around this difficulty by arguing that such overlaps occurred “at irregular intervals.” Somehow, the crossbills maintained their distance from one another, not exchanging genes, although at times they might be feeding in the same trees in the same forests. How they kept separate when sharing nesting habitats remained unclear.
In the 80 years since, other investigators have proposed other subspecies, many of them in conflict with Griscom’s original scheme. Alternate schemes and terminology have also been proposed: “pseudo-species,” “morpho-types,” and others. No consensus has emerged, except that Red Crossbills are even more difficult to understand than Griscom realized.
One distinctive population unknown to Griscom was discovered in 1912 by A. C. Bent on a visit to Fox Island River, Newfoundland. Studying two specimens collected by his host in Canada, he noted their large, heavy bills and several other differences between them and the eastern Red Crossbills in New England. He collected about a dozen other individuals and typed up an introduction to a report intended to describe a new subspecies—the “Newfoundland crossbill,” L. c. percna. The report was not completed, however. It was found in his papers after his death in 1954 and was not published until 1968.
Today, the Newfoundland population seems to be one group that behaves as a subspecies is supposed to: it remains isolated in its homeland island and, as far as we know, has not bred elsewhere. It may also be the most endangered population, with numbers in a steep decline since red squirrels were introduced to the island in the 1960s. Apparently, the squirrels outcompete the crossbills for conifer seeds. Logging has also taken a toll. A 2004 report by the Committee on the Status of Endangered Wildlife in Canada estimated that only 500 to 1,500 Newfoundland crossbills survive.
The ornithologist whose work on Red Crossbills may be most familiar to birders is J. G. Groth, who conducted a series of studies in the late 1980s and early ’90s that led him to designate eight populations, many of them overlapping with one another, sometimes breeding close to one another, and each identifiable by its flight call. This work is the source of the nine flight calls described in The Sibley Guide to Birds and elsewhere. Some of Groth’s call types match populations identified by earlier researchers (the type 8 call is given by the Newfoundland birds, for example), but others do not. And the calls sound similar to most of us. I once met a birder in the New Jersey Pine Barrens who pointed in the direction of a small bird that had just passed overhead and said, “Did you hear that? Red Crossbill—type 4!” I almost countered, “Are you sure? I had it in my bins for at least two seconds at 100 yards, and the bill curvature looked more like a type 3.” Instead, I bit my skeptic’s tongue. Birders like that are pushing the frontiers of identification, and we should wish them good fortune.
Groth himself concluded that the Red Crossbill populations are “cryptic species”—a term used most often by entomologists to describe insect populations whose members can distinguish their sexual partners from look-alikes, although human observers cannot—without using microscopes and chemical analyses.
Whether Groth’s view will eventually prevail, only further research will determine. In the meantime, birders can note that we are spoiled by the luck of our draw. With the apparent exception of the Red Crossbills and perhaps a handful of other species in North America, our favorite creatures see and hear each other and distinguish each other just as we do. No law of nature requires that situation, and entomologists, botanists, and lichenologists, who encounter impossible identification problems far more often, envy us and our usual certainty.
All About Birds
is a free resource
Available for everyone,
funded by donors like you
American Kestrel by Blair Dudeck / Macaulay Library